Rosseto, V. & Santos, F.A.M. 2006.
A heterogeneidade ambiental de três fisionomias florestais em um fragmento em Campinas (SP).
CD de Resumos do 57º Congresso Nacional de Botânica, Resumo 3285.
Resumo expandido:
Introdução
A heterogeneidade ambiental se refere à distribuição descontínua dos fatores abióticos e bióticos ao longo do espaço [1], e tal variação é dependente da escala de estudo [2]. Na escala micro-local, a heterogeneidade ambiental está associada com a variação espacial de micro-sítios adequados para a germinação e o estabelecimento de espécies vegetais [3]. Com isso, supõe-se que a distribuição de espécies arbóreas tropicais esteja relacionada com a distribuição heterogênea dos fatores ambientais [4, 5]. A heterogeneidade pode ser avaliada em termos de descritores ecológicos e pode ser medida de forma quantitativa e/ou qualitativa [2]. Com o objetivo de se avaliar a heterogeneidade ambiental em três fisionomias florestais na área de estudo foram propostas as seguintes questões: (1) Qual a variabilidade espacial da abertura do dossel, da umidade do solo, da profundidade de serrapilheira e da abundância de árvores em três fisionomias florestais em diferentes escalas? (2) Qual a relação entre os fatores quantitativos analisados na escala micro-local?
Material e métodos
A. Área de estudo
O estudo foi realizado na Área de Relevante Interesse Ecológico (ARIE) de Santa Genebra (22°49'S e 47°06'W) [6], maior fragmento urbano do município de Campinas (SP), com 251,77 ha [7]. A maior parte da ARIE é composta por floresta estacional semidecidual, mas há áreas descontínuas de floresta paludosa dentro do fragmento. Além disso, cerca de 0,5 ha da ARIE apresenta o dossel composto quase predominantemente por apenas uma espécie, maria-preta (Diatenopteryx sorbifolia Radlk), com sub-bosque ralo e presença de uma série de espécies restritas apenas a este local. Portanto, devido as suas peculiaridades, tal região foi denominada de maciço de maria-preta [8].
B. Coleta de dados
Para o estudo foram consideradas três fisionomias florestais presentes na ARIE: floresta estacional semidecidual, denominada no estudo apenas como floresta (FL); floresta paludosa (FP); e maciço de maria-preta (MP). Em cada fisionomia foram alocadas três amostras. Cada amostra foi composta por 40 parcelas de 5 x 5 m, totalizando 0,3 ha em cada fisionomia estudada (360 parcelas no total). Para a avaliação da abertura do dossel foi tirada uma foto hemisférica a 1,30 m de altura do solo em cada parcela, sendo as fotos posteriormente analisadas pelo programa Gap Light Analyser versão 2.0 [9]. Para a umidade do solo foi coletada uma amostra por parcela a 20 cm de profundidade, sendo avaliada a razão: (peso solo úmido – peso solo seco)/(peso solo úmido). A profundidade de serrapilheira foi medida em quatro pontos em cada parcela, com o auxilio de uma régua de metal graduada, sendo usada a profundidade média por parcela. A abundância de árvores foi avaliada pela densidade de árvores com PAP (perímetro à altura do peito) ³ 15 cm em cada parcela.
C. Análises estatísticas
Para as análises foram consideradas três escalas: escala local (fisionomias), escala intermediária (amostras) e escala micro-local (parcelas). As variáveis foram comparadas por “box-plots” no programa SYSTAT versão 10.2 [10]. Tais gráficos mostram o intervalo de confiança da mediana, de modo que se pode detectar diferenças significativas entre os grupos [11]. Para verificar se os dados apresentavam autocorrelação espacial na escala micro-local foram feitos correlogramas, que são gráficos cujos coeficientes de correlação espacial - coeficiente Z de Moran padronizado - são colocados na ordenada em função da distância entre as localidades [12, 13]. Para testar a significância dos correlogramas foi utilizada a correção de Bonferroni para testes múltiplos [12]. A correlação entre as variáveis na escala micro-local foi testada através do teste parcial de Mantel, que considera variáveis com autocorrelação espacial [12, 13]. Tanto os correlogramas, como as correlações foram feitos no programa Passage versão 1.1 [13].
Resultados
A. Escalas local e intermediária
A abertura do dossel foi menor do que 10% em todas as fisionomias, com a floresta paludosa apresentando maiores valores, enquanto que a floresta e o maciço de maria-preta não diferiram (Fig. 1A). Na escala intermediária observou-se variação na abertura do dossel entre as parcelas, em especial na floresta paludosa (Fig. 1A). A umidade do solo foi maior na floresta paludosa, seguida da floresta e do maciço de maria-preta (Fig. 1B). Na escala intermediária verificou-se grande variação de umidade do solo entre amostras na floresta paludosa, com uma das amostras (FP3) apresentando valores de umidade semelhantes às outras fisionomias (Fig. 1B). A profundidade de serrapilheira na escala local foi similar entre as áreas de floresta e do maciço de maria-preta, enquanto que na floresta paludosa verificou-se uma tendência de menor profundidade da camada de serrapilheira (Fig. 1C). Na escala local, o número de árvores com PAP > 15 cm foi maior na floresta paludosa com 745 indivíduos, seguida do maciço de maria-preta com 442 indivíduos e floresta com 429 indivíduos. Na escala intermediária, pelo menos uma das amostras em todas as fisionomias analisadas diferiu na densidade de árvores das outras amostras dentro de uma mesma fisionomia (Fig. 1D).
B. Escala micro-local
A abertura do dossel na escala micro-local, em geral, apresentou pequena variabilidade entre as parcelas em todas as fisionomias, sendo que a maior parte das amostras não exibiu autocorrelação espacial (Fig. 2A). Por outro lado, a maioria das amostras apresentou autocorrelação espacial para a umidade do solo, destacando-se as amostras da floresta e da floresta paludosa, que apresentaram correlogramas com formato parecido (Fig. 2B). A profundidade de serrapilheira também apresentou autocorrelação espacial para a maioria das amostras (Fig. 2C). No caso da abundância de árvores por parcela, verificou-se que a maioria das amostras apresentou padrão aleatório, com exceção de duas amostras na floresta paludosa (Fig. 2D). Com relação às correlações, a maioria não foi significativa. As exceções ocorreram em duas amostras distintas do maciço de maria-preta, onde a abertura do dossel foi correlacionada negativamente com a densidade de árvores na MP2 (-0,11; p=0,040) e com a umidade do solo na MP3 (-0,15; p=0,032). Já em uma amostra de floresta paludosa (FP2), a abertura do dossel foi correlacionada positivamente com a densidade de árvores (0,34; p<0,000). A umidade do solo foi correlacionada negativamente com a profundidade de serrapilheira em uma repetição da floresta paludosa (FP2) (-0,12; p=0,049). A profundidade de serrapilheira foi correlacionada positivamente com o número de árvores em uma amostra da floresta paludosa (FP1) (0,22; p=0,016).
Discussão
A floresta paludosa exibiu maior abertura do dossel do que as outras fisionomias florestais, assim como encontrado em outros estudos [14, 15]. Na escala micro-local, a ausência de autocorrelação espacial a distâncias menores do que 40 m indica que a luz apresenta distribuição espacial imprevisível, devido à composição das espécies [14], composição e estrutura dos estratos inferiores [16] e variabilidade na distribuição e tamanho das clareiras [4]. No caso das variações da umidade do solo em florestas paludosas em escala intermediária, estas podem estar relacionadas com o nível e afloramento do lençol freático, calha, volume e velocidade da água [17]. As características físicas do solo no maciço de maria-preta, como baixa umidade e solo muito compacto, além da baixa densidade de raízes a 20 cm de profundidade do solo, pode ser um indicativo de que o solo seja um fator restritivo para o estabelecimento das espécies arbóreas neste local. Para a profundidade de serrapilheira, outros estudos indicam que esta pode variar em escalas menores do que um metro [18]. Contudo, encontrou-se agregação espacial em pequenas escalas, que pode estar relacionada com a composição das espécies, em especial as decíduas [14] e presença de árvores e /ou galhos caídos recentemente. O padrão aleatório das árvores em escala micro-local pode ser o resultado da grande heterogeneidade das condições ambientais, gerando assim tal padrão para a comunidade. Já a presença de autocorrelação espacial nas primeiras classes de distância para árvores na floresta paludosa pode estar associada com variações na microtopografia do solo, pois em áreas mais elevadas, não alagadas, há maior densidade de árvores. Além disso, algumas espécies ocorrem restritamente nestas elevações, indicando especialização de habitat em escala micro-local [15, 19]. A correlação negativa entre abertura do dossel e umidade do solo, e abertura do dossel e abundância de árvores no maciço de maria-preta pode estar relacionada com a estrutura do sub-bosque, que é composto por poucas espécies e estrato herbáceo ralo. Com isso, sugere-se que em locais onde os estratos inferiores sejam menos densos, haja maior correlação entre a abertura do dossel medida a 1,30 m e a profundidade de solo avaliada, e menor a influência de tais estratos sobre a disponibilidade de luz. A correlação positiva entre a abundância de árvores e abertura do dossel, pode estar relacionada com a composição da comunidade na repetição da floresta paludosa (FP2). Neste local, o dossel é composto basicamente por Euterpe edulis Mart. e devido a arquitetura e o formato das folhas desta espécie, o dossel apresenta zonas extensas de luz entre os palmitos, diferente de um dossel composto por espécies dicotiledôneas. Considerando-se as variáveis analisadas, verificou-se que a umidade do solo foi o fator mais determinante na diferenciação entre as fisionomias (escala local), enquanto que na escala intermediária em geral observa-se ampla variação de todos os fatores estudados, dentro de todas as fisionomias avaliadas. Na escala micro-local, a grande variação quantitativa de condições ambientais encontradas fornece indicativos sobre o elevado número de combinações possíveis entre as variáveis ambientais, que influenciam a regeneração das espécies arbóreas tropicais.
Referências
1. DALE, M.R.T. 1999. Spatial pattern analysis in plant ecology. New York, Cambridge University Press. 326p.
2. LI, H. & REYNOLDS, J.F. 1995. On definition and quantification of heterogeneity. Oikos, 73: 280-284.
3. HUTCHINGS, M.J. 1989. The structure of plant populations. In: CRAWLEY, M.J. (Ed.). Plant Ecology. Oxford, Oxford Blackwell. p. 97-136.
4. DESLOW, J.S. 1980. Gap partitioning among tropical rainforest trees. Biotropica, 12 (suppl.): 47-55.
5. RICKLEFS, R.E. 1977. Environmental heterogeneity and plant species diversity: a hypothesis. American Naturalist, 111: 376-381.
6. DECRETO n°91.885 de 05/11/1985. Diário Oficial da União, Brasília.
7. LEITÃO-FILHO, H.F. 1995. A vegetação da reserva de Santa Genebra. In: MORELLATO, P. C. & LEITÃO-FILHO, H. F. (Eds). Ecologia e preservação de uma floresta tropical urbana: Reserva de Santa Genebra. Campinas, Editora da Unicamp. p. 97-136.
8. NAVE, A.G. 1999. Determinação de unidades ecológicas num fragmento de floresta nativa, com auxílio de sensoriamento remoto. Dissertação de Mestrado, ESALQ, Piracicaba.
9. FRAZER, G.W., CANHAM, C.D. & LERTZMAN, K.P. 1999. Gap Light Analyzer (GLA). Imaging software extract canopy structure and gap light transmission indices from true-colour fisheye photographs, users manual and program documentation. Version 2.0. New York, Simon Fraser University, Burnaby, British Columbia, and The Institute of Ecosystem Studies, Millbrook.
10. SPSS, INC. 2000. SYSTAT version 10.2, standard version. SPSS, Inc. Chicago.
11. MC GILL, R., TUKEY, J.W. & LARSEN, W.A. 1978 Variations of Box Plots. The American Statistician, 32: 12-16.
12. LEGENDRE, P. & FORTIN, M-J. 1989. Spatial pattern and ecological analysis. Vegetatio, 80: 107-138.
13. ROSENBERG, M.S. 2001. Passage. Pattern analysis, spatial statistics, and geographic Exegesis. Version 1.1. Department of Biology, Arizona. State University, Tempe. AZ.
14. BIANCHINI, E., PIMENTA, J.A. & SANTOS, F.A.M. 2001 Spatial and temporal variation in the canopy cover in a tropical semi-deciduos forest. Brazilian Archives of Biology and Technology, 44: 269-276.
15. SOUZA, A.F. & MARTINS, F.R. 2005. Spatial variation and dynamics of flooding, canopy openness, and structure in a Neotropical swamp forest. Plant Ecology, 180:161-173.
16. NICOTRA, A.B.; CHAZDON, R.L & IRIARTE, S.V.B 1999. Spatial heterogeneity of light and wood seedling regeneration in tropical wet forests. Ecology, 80: 1908-1926.
17. TONIATO, M.T.Z., LEITÃO-FILHO, H.F. & RODRIGUES, R.R. 1998. Fitossociologia de um remanescente de floresta higrófila (mata de brejo) em Campinas, SP. Revista Brasileira de Botânica,21: 197-210.
18. MOLOFSKY, J. & AUGSPURGER, C.K. 1992. The effect of leaf litter on early seedling establishment in a tropical forest. Ecology, 73: 68-77.
19. SOUZA, A.F. & MARTINS, F.R. 2004. Microsite specialization and spatial distribution of Geonoma brevispatha, a clonal palm in southeastern Brazil. Ecological Research,19: 521-532.
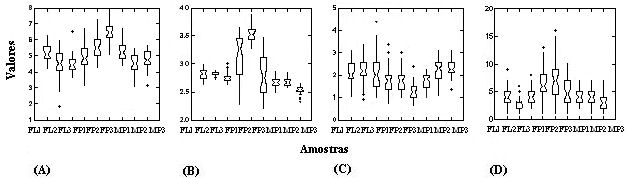
Figura 1. Distribuição das medidas quantitativas avaliadas no estudo, indicando a variação entre as fisionomias, entre as amostras e entre as parcelas. (A) porcentagem de abertura do dossel, (B) logaritmo da porcentagem de umidade do solo, (C) profundidade de serrapilheira (cm) e (D) densidade de árvores por parcela. Dados da porcentagem de umidade do solo foram transformados em log para diminuir a grande amplitude de variação dos dados. FLi, FPi e MPi correspondem a amostras da floresta, da floresta paludosa e do maciço de maria-preta, respectivamente. Sobreposição entre os intervalos de confiança nos “box-plots” indicam igualdade entre as amostras.
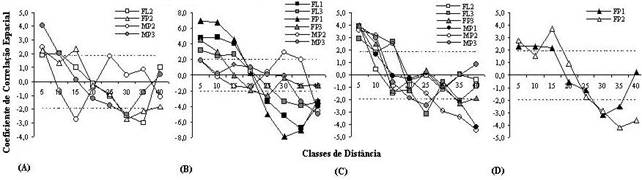
Figura 2. Autocorrelação espacial das variáveis estudadas nas amostras, indicadas pelos correlogramas. São mostrados apenas os correlogramas significativos. (A) abertura do dossel (%), (B) umidade do solo (%), (C) profundidade de serrapilheira (cm) e (D) densidade de árvores FLi, FPi e MPi correspondem a amostras da floresta, da floresta paludosa e do maciço de maria-preta, respectivamente.Valores entre linhas tracejadas indicam padrão espacial aleatório para classes de distância individuais, segundo o coeficiente Z (coeficiente I de Moran padronizado).