Species and Interaction
Diversity in Plants and Phytophagous
Insects
Principal
investigator: Thomas Michael Lewinsohn
<thomasl@unicamp.br>omasl@unicamp.br
Laboratório de Interações
Insetos-Plantas
Instituto de Biologia, Unicamp
13081-970 Campinas, SP, Brasil
Phone +55-19-7887022 Fax +55-19-2893124
PROJECT
INTRODUCTION
Most
inventories and censuses of biodiversity
in terrestrial systems are conducted with
vascular plants and vertebrates, and
directed at particular taxa. Considerable
expertise and substantial data sets are
available for plants (especially
angiosperms) and various classes of
vertebrates for most habitats and
ecoregions on the planet. However, the
bulk of the world’s biodiversity
belongs to other taxa, notably
arthropods, which are far less known and
surveyed. In recent years, many authors
have stressed the need to increase and
organize efforts of surveying,
inventorying, describing arthropods and
other taxa. This entails a major
capacity-building effort in taxonomy as
well as the development of new methods to
obtain samples of various taxa in
different ecological settings, as well as
new procedures to handle and process
these samples and to analyse the ensuing
data.
Arthropods
offer a number of advantages for
inventorying or monitoring diversity
(Kremen et al., 1993): they occur in
virtually any terrestrial habitat; their
distribution, geographical and among
habitats, is often finer-grained than
that of vertebrates and they respond
readily to a number of ecological
processes; thus, they are potentially
highly suitable as indicators of
geographical or ecological uniqueness or
to give early warning of various forms of
disturbance. Moreover, their small body
sizes and often large numbers make mass
collecting feasible which, together with
high species numbers, in turn make them
suitable for statistical assessment of
total species richness and its breakdown
in various ways, notably into local and
regional components and their turnover
(e.g. Lewinsohn, 1991; Colwell and
Coddington, 1994).
Inventories
of arthropods will often concern whole
taxa, at the family level or higher
(Coddington et al., 1991) but recent
proposals have advocated the advantages
of more restricted sampling, in two
different ways: either by concentrating
on certain well-circumscribed habitats,
such as single tree canopies sampled by
fogging (e.g. Basset, 1996) or by the
restriction to a smaller,
"ecologically informative"
taxon, for instance a single butterfly
genus (Kremen, 1994).
In the
present project we intend to build on our
previous experience in investigating the
diversity of a set of terrestrial insects
defined by a common resource set rather
than by a purely taxonomic criterion
(Lewinsohn, 1991; Müller-Schärer et
al., 1991; Lawton et al., 1993). Since
plants are the resource base on which we
survey and record phytophagous insects,
plant diversity is both an objective in
itself and a potential determining factor
for the diversity of insects associated
to these plants.
Our
approach is somewhat similar to studies
of canopy arthropod assemblages. However,
we concentrate on endophagous insects
that are collected within plants and
reared out from them. Thus, while the
variety of taxa and the size of the
assemblages we study are much smaller
than that encountered by external
collecting methods such as canopy
fogging, our approach ensures that every
insect is feeding in the plant and that
they are indeed trophically linked. Two
consequences follow on that. First, such
species assemblages are parts of food
webs and therefore can be examined and
evaluated for food-web patterns and
traits. We are particularly concerned,
for instance, with the degree of
compartmentation of local assemblages,
which should affect their stability and
resilience to disturbance or their
response to new additions, be it
introduced plants or insects (Pimm, 1991;
Polis and Winemiller, 1996). Beyond the
number and species makeup of a local
biotic assemblage, its structure of
trophic associations – its
interaction diversity (Thompson, 1997)
– is potentially useful as indicator
of biological diversity and to monitor
responses its changes.
The second
consequence of targeting assemblages of
endophagous insects as a study system is
their evolutionary relationship to host
plants. Host associations evince
long-term, phylogenetic patterns (Farrell
et al., 1992) as well as short-term
adjustments to host distributions and
combinations and their variation in time
and space (Thompson, 1994). Thus,
endophagous insect diversity can be
studied for their phylogenetic structure
and constraints, but also as to their
short-term response to shifting host
conditions and combinations among
localities and in time. In this regard,
intraspecific genetic differentiation
among host plants or among localities is
an important question which we intend to
address. Genetic studies of congeneric
phytophagous species are essential for an
assessment of the importance of
phylogenetic restraints and radiation in
establishing them (or not) on different
hosts. Furthermore, studies of genetic
similarity of the host plants themselves,
both among conspecific populations and
among species, provide further insights
into the evolutionary establishment and
maintenance of insect-plant associations.
The
approach which we have been developing in
previous work and which we propose for
the current project, allows us to address
biological diversity simultaneously at
the level of species diversity and
genetic diversity. Diversity at these two
levels of organization is evaluated at
different spatial scales, from the local
host plant patch up and across different
habitats and ecoregions. Local and
regional components of biological
diversity and their organization and
dynamics at these different scales, are
one of the main concerns of current
ecological research (e.g. Lewinsohn,
1991; Ricklefs and Schluter, 1993;
Ricklefs et al., 1995).
Among
other potential correlates and predictors
of insect biological diversity we single
out the secondary chemistry of their host
plants. Chemical diversification of
plants is intimately related to insect
diversification and a major candidate to
explain both long-term, phylogenetically
structured patterns of association, and
short-term preferences or host shifts in
local assemblages (e.g. Cates, 1980).
Chemical screening of plants is one of
the main thrusts of biodiversity
prospecting, but usually with little
concern for the ecological implications
of chemical diversity. The Asteraceae, on
which our current work is targeted, are
well-known for their high chemical
diversity; however, most neotropical
species have never been investigated for
their secondary chemistry. Thus, chemical
diversification of the Asteraceae is an
important potential explanatory factor
for the diversity and patterns of
association within our target
assemblages; but the chemical survey of
plants within this family is a goal of
interest in itself.
ASTERACEAE
AND FLOWERHEAD INSECTS AS A STUDY SYSTEM
The main
system for this project are the
Asteraceae (Compositae) and their
associated phytophagous insects,
especially the flowerhead feeding
insects, in Southern and Southeast
Brazil. Asteraceae fulfill several
prerequisites and offer a number of
advantages:
- nearly
ubiquitous, with a rich gamut of
wide-ranging species up to strict
habitat endemics
- cosmopolitan,
allowing comparisons across
localities up to different
continents and life zones
- taxonomically
well studied; major subdivisions,
especially tribes, fairly
well-established, with proposed
phylogenies for many major
sections within the family
(Bremer, 1994)
- they
include serious weeds extensively
canvassed for biocontrol agents;
in South America, these include Chromolaena
odorata (Crutwell, 1974), Baccharis,
and several Heliantheae (Daniel
Gandolfo, USDA/Argentina,
unpublished).
- they
also include several
"classical" systems for
the study of the ecology and
evolution of insect-plant
interactions, such as Senecio,
Solidago, Centaurea,
Haplopappus, their herbivores
and parasitoids (Louda, 1982;
Zwölfer, 1988; Root and
Capuccino, 1993; Abrahamson and
Weis, 1997).
Most
endophagous insects associated with
Asteraceae flowerheads are restricted to
this family, and several (e.g. Diptera
Tephritidae, Agromyzidae and
Cecidomyiidae; Lepidoptera Pterophoridae
and Tortricidae) count Asteraceae among
their major, if not the foremost, host
group. Thus, these insect taxa have
evolved in close association with
Asteraceae (Zwölfer, 1988; Lewinsohn,
1991).
Other
study systems may be included in latter
phases of the project, to validate
results from the Asteraceae –
flowerhead insect system.
PREVIOUS RESULTS
Results
from previous grants are summarized in a
later section (p.*). Here we give a brief
history of this line of investigation
within our research group.
The
Asteraceae – flowerhead insect
system has been studied by us since 1985.
Initial samples established the general
pattern of associations and partitioned
local and regional diversity in various
habitats of Southeast Brazil (Lewinsohn,
1991). Further study has concentrated on
a local assemblage of Vernonieae and
Tephritidae (Prado and Lewinsohn, 1994;
ms in prep) and the genetics of Tomoplagia,
the main tephritid genus in this
assemblage, has also been studied in
detail, showing a close match of host
plant and genetic similarities (Solferini
et al., in prep).
In an
ensuing project from 1995 to 97 we
collected and processed 1510 samples of
more than 400 species of Asteraceae (over
20% of known species in Brazil) in two
montane areas: the southern Espinhaço
range in the State of Minas Gerais, and
the coastal ranges and highlands in Santa
Catarina and Rio Grande do Sul. Almost 17
000 insects were reared, sorted and
(except for gall midges) mounted from
these samples. Till now, Tephritidae,
Tortricidae, Pterophoridae and Pyralidae
have been wholly sorted, and Agromyzidae
are near completion.
In all,
167 plant species were sampled at least
three times in at least two localities,
and 65 species have been sampled in five
or more localities, thus being adequate
for an analysis of local and regional
richness. The whole database provides an
extensive matrix of plants, insects and
their association across various
localities. This represents a unique
resource for analyzing the distribution
of diversity within and across
localities, regions, and host taxa of
different levels.
Genetic
studies are being completed on two
tephritid genera: Tomoplagia and Xanthaciura
and also on two endemic plant genera.
Chemical studies have investigated the
structure and interspecific diversity and
similarity of pyrrolizidine alkaloids
within two genera, Senecio and Trichogonia
(sensu lato, including Trichogoniopsis
Robinson). Further studies are being
directed at other genera of the
Eupatorieae, particularly in the Eupatorium
complex itself.
CERRADO
Conservation
of the Cerrado biome has been of less
concern than Amazonian and Atlantic rain
forest or the Pantanal; yet it is under
immediate threat, and disappearing at a
faster rate than any other major
terrestrial biome in Brazil. Cerrado is
also under-represented in conservation
areas. This situation was recently
revised at the national level
(PROBIO-MMA, 1998) and a synthesis of
proposed priorities for conservation
within the biome is forthcoming. Within
São Paulo, a major study integrating
existing information has resulted in a
comprehensive survey of existing Cerrado
areas and remnants and a proposal for
priorities in their conservation and
management (São Paulo - SMA, 1997).
Given the
current effort in improving conservation
and management efforts within the Cerrado
region, the considerable information
(especially floristic) already available,
and the fair representation of
Asteraceae, especially among herbs and
shrubs, the cerrado is a sensible choice
for a major target of this study, noting
also that the cerrado biome does include
the highland fields and montane meadows
(campos rupestres) which are among the
richest and most diverse areas in
Asteraceae in Brazil, and which have been
the main study areas of our recent work
on this system.
OBJECTIVES
We propose
to:
- Investigate
the structure of biological
diversity in subsets of natural
communities composed of plants
and their associated phytophagous
insects, using Asteraceae and
flowerhead insects as a base
system.
- Sample
Asteraceae and their associated
phytophages in extensive areas
and remnants of cerrado in the
State of São Paulo and, for
comparative purposes, in cerrado
core areas in adjoining states.
- Develop
or experiment existing methods to
separate total biological
diversity in components of
different spatial, temporal and
structural scales, e.g.
within-host diversity and among
host turnover, and local
diversity versus species
richness.
- Correlate
diversity of Asteraceae and their
endophages with that of other
taxa or community subunits in
cerrado areas.
- Extend
the existing database on
Asteraceae and phytophagous
insects by sampling other areas,
especially the Mantiqueira range
on the northeastern border of
São Paulo; its transition into
the Espinhaço range in Southern
Minas Gerais; and various open
habitats in montane and coastal
areas within the Atlantic
rainforest biome.
- Investigate
the contribution of genetical,
phylogenetic, chemical, and
ecological factors in determining
species diversity or interaction
diversity and its variation on
different spatial, structural and
time scales.
- Investigate
the response of species
diversity, interaction diversity
and species composition to
various regimes or histories of
disturbance or impact, through
comparisons of assemblages in
preserved Cerrado areas with
those in nearby sites.
- Investigate
the structure of species
interactions in these systems by
applying existing analyses and
theory or by developing new
approaches.
RATIONALE
The aims
of this project are in accordance with
the objectives of the Special Research
Program "Conservation and
sustainable use of the biodiversity of
the State of São Paulo - BIOTA/SP"
(see p.*).
Within the
general goals of the BIOTA-SP initiative,
the present proposal fits in most clearly
with the following: (1) "to study
and know the biodiversity of the State of
São Paulo and to disseminate this
knowledge and its importance"; (2)
"to understand the processes that
generate, maintain and impact this
biodiversity"; (5) "to develop
methodological bases and reference
standards for studies of environmental
impact." We expect this project to
contribute especially to goals 2 and 5.
The
proposed study may also help to attain
several other goals of this initiative,
but we deem goal 13 to be exceptionally
important: "to contribute to implant
the Convention of Biological Diversity in
the State of São Paulo, joining national
and international efforts for the
knowledge, conservation and sustainable
use of biodiversity". Our proposed
effort to investigate biodiversity
jointly at the genetic and the species
level and at different spatial scales so
that results may be interpretable up to
the landscape level as well, may
represent its most evident contribution
to attain this general goal.
Prioritization
of the Cerrado biome among the several
ecosystems to be studied has been
discussed in the Introduction. This
should also fit other projects concerning
Cerrado in the BIOTA-SP initiative (such
as proposals 13 and 17).
METHODS
FIELD
SAMPLING AND REARING
Our basic
information unit is the use of a plant
species by an insect species in a given
locality, derived from our field records
and samples. Thus, the project will
produce biogeographical information on
Asteraceae and their endophagous insects,
and on the associations among them as
well. The minimal information set for our
records comprises the locality name,
GPS-recorded coordinates, altitude,
habitat, collector name, and date. These
data will be made available to the
Biodiversity Data Base of the State of
São Paulo, and to that end they conform
to the minimum standards established for
all projects under the BIOTA-SP
initiative.
The
general sampling procedure within a
chosen locality consists of:
(a)
surveying Asteraceae at the site
and recording their location and
topographic or habitat
preference. In extensive field
trips, no quantitative assessment
of plant populations is made.(b)
selecting species to be sampled
among those in flower, according
to the objectives of that field
trip.
(c)
collecting flowerheads of
selected populations. Every plant
sample is documented with voucher
specimens and a GPS fix.
Additional information on the
plant and the site, such as
habitat type, phenology and plant
population size, is also
recorded. Whenever necessary,
photographs of plant habit and
details of flowering branches and
flowerheads are made as mnemonic
aids and to help sorting of more
difficult species groups.
(d)
collecting plant samples into
appropriate solvents (usually,
pure ethyl alcohol) for chemical
analysis.
In the
laboratory, flowerheads are maintained in
containers with a meshed cover to rear
out adult insects. Samples are inspected
periodically and emerged adults are
stunned with a stream of CO2 ,
stored in vials and later mounted on
entomological pins, except for
cecydomyiid flies, which are preserved in
alcohol or will be mounted in microscope
slides (Gagné, 1994). After adult
emergence ceases, within 60 days after
collecting, samples are dried and
weighed. A fraction of flowerheads from
each sample is counted, dried and weighed
to establish individual flowerhead
weights, used to estimate total numbers
of flowerheads in samples from their dry
bulk weight.
SORTING
AND IDENTIFICATION
Mounted
specimens are sorted into morphospecies
based on usual characters for each group;
in some cases, e.g. Agromyzidae, this
entails dissection and mounting of
genitalia of specimens from each sample.
Sorting is aided by reference collections
("morphotypes") and by
photographs and photomicrographs of whole
specimens (habitus) and relevant
details.
Identifications
are based on specimens previously
identified by specialists and on
published descriptions. Plant
identifications are checked by
participating and collaborating
investigators (João Semir, Roberto
Esteves, Nicholas Hind, John Pruski;
others to be added). Subsets of reared
insects are sent to collaborating
investigators for identification or
verification (Allan Norrbom, Diptera
Tephritidae; Vitor Becker, Lepidoptera
Tortricidae; Cees Gielis, Lepidoptera
Pterophoridae) and occasionally to other
taxonomists.
The only
major group of flowerhead endophages for
which we have no established procedure as
yet are Diptera Cecidomyiidae. Genera
commonly found in asteraceaous
flowerheads can be identified fairly
readily according to Gagné (1994 and
references therein). However,
identification at the species level is
demanding (Raymond Gagné, personal
communication) and we may presume that
our samples include a fair number of
undescribed taxa. Therefore, to saort
these midges reared specimens and pupal
cases from each sample will be mounted on
microscope slides and then examined under
interference microscopy, according to
Gagné (1994 and personal communication).
We intend to set up a reference
collection of microscope slides and
digitize pictures of adults (with special
attention to antennal segments, legs, and
genitalia) and larvae, especially the
sternal spatula. These often present
usable characters in this family, and
discrete characters may offer a basis for
an operational separation of
morphospecies in the group. After the
first year, we will evaluate our results
and the effort involved, to decide
whether we will continue to attempt to
separate the family to species level.
Insect
specimens will be stored in reference
collections in our lab and in the
permanent collections of the Museu de
História Natural of Unicamp. Type
specimens will be placed in the Museu de
Zoologia of USP (MZUSP), São Paulo.
Plant specimens will be stored in the
Unicamp Herbarium. Selected specimens may
be lent or donated to other collections
in Brazil or overseas.
DATA
STORAGE AND ANALYSIS
Both in
field work and in rearing and sorting, we
intend to use sample identification and
data entry with barcoding, to reduce
errors and improve efficiency. We also
intend to use digital images of sampled
plants and insects as aids to recognition
and identification, incorporating them in
the project data base. One of the
intended products of the project is a
visual identification system and guide to
the organisms in this study, both plants
and insects.
GENETIC
STUDIES AND PHYLOGENETIC ANALYSES
Genetic
studies focus on phytophagous genera
comprising several species with
well-established host ranges, varying
from strict monophagy to several tribes
of Asteraceae. Previous studies have
concentrated on two genera of tephritids.
Within this project, Tephritidae will
still be the main study group but others
will be added, starting with the genus Melanagromyza
(Diptera Agromyzidae).
Genetic
variability of populations will be
evaluated through several analytical
techniques, using nuclear and cytoplasm
markers in combination with the
sequencing of particular regions to
produce phylogenies. Markers with
different modes of inheritance allow
better and more detailed inferences on
observed patterns. Eletrophoretic
analysis of isozymes, the main technique
employed in previous work, will be used
as a complementary technique to examine
the extent of genetic differentiation
among hosts and/or localities, and
whether variability covaries with host
range.
Variability
and interspecific differentiation of host
plants will also be examined through
enzyme and DNA studies, concentrating at
first on populations of endemic and
restricted species and genera.
SECONDARY
CHEMISTRY OF ASTERACEAE
Following
on previous work, we intend to screen
host plants for pyrrolizidine alkaloids.
Work will span plants in various tribes
but concentrate on Eupatorieae and
Senecioneae. Fresh flowerheads are
collected in ethanol and acid-base
extracts are analyzed through gas
chromatography - mass spectrometry. Other
chemical groups will be added to the
screening program if feasible.
SAMPLING PLAN
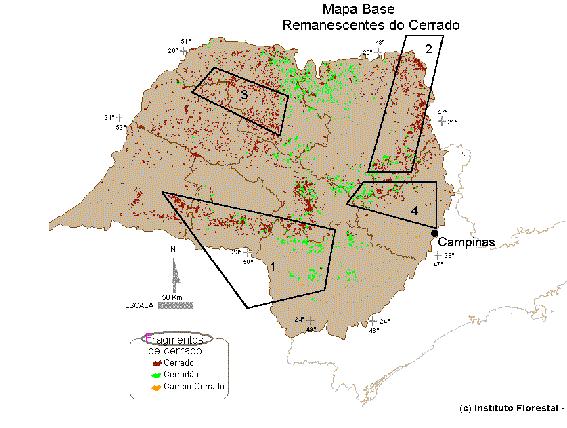
CERRADO
STUDY
In project
years 1-2, ten representative localities
of Cerrados will be chosen within São
Paulo according to their size,
preservation status, plant species
richness and distance from others sites.
Each site will be sampled once at the
beginning of the dry season and once in
the rainy season, during two years. Five
sites within the core area of the
Cerrados (Minas Gerais, Goiás and
Distrito Federal) will be sampled twice
within a year, to compare their diversity
and composition to the São Paulo sites,
which are fairly close to the southern
limit of cerrado distribution. These
initial samples will serve to establish
the general insect-plant association
matrix for assemblages within the
cerrado.
In years
2-4 of the project, a larger number of
sites (30 or more) will be sampled,
concentrating on commoner and widespread
plant species (ca. 10 species). These
sites will include areas with different
kinds of use and, preferably, with a
known history. They will serve to tease
out the relationship between local and
regional insect species richness and the
response of insect assemblages to
different forms of use.
Within the
cerrado biome, further montane areas will
be sampled to complete the insect-plant
array, in one or two extended field trips
per site.
MANTIQUEIRA
AND OTHER AREAS
Asteraceae
in the Serra da Mantiqueira and its
transition into the Espinhaço will be
sampled in years 1-3 in three different
seasons: early wet season, late wet
season, and early dry season. Mantiqueira
sites will spread out from two foci:
Campos do Jordão (SP) and Itatiaia (RJ).
The transition to the Espinhaço will be
sampled in a broad west-east arch from
the Serra da Canastra up to the Parque do
Ibitipoca (MG).
Montane
and coastal areas within the Atlantic
forest region will be sampled in two
field trips per site. Sites will coincide
if feasible with those of other projects
in the BIOTA-SP initiative.
PERMANENT
SITES
We intend
to establish at least one permanent site
to initiate a longer-term study of host
populations and their associated insects,
to be surveyed extensively and then
monitored for at least five years. Since
Unicamp has no field station, choice of
site(s) will depend on their quality,
guarantees that the study area will not
be cleared or disturbed, and
authorization to conduct field
experiments.
LITERATURE CITED
Abrahamson, W.G. and
Weis, A.E. 1997. Evolutionary
ecology across three trophic
levels: goldenrods, gallmakers,
and natural enemies. Princeton
University Press, Princeton, New
Jersey, 456 pp.Basset, Y.
1996. Local communities of
arboreal herbivores in Papua New
Guinea: predictors of insect
variables. Ecology, 77:
1906-1919.
Bremer,
K. 1994. Asteraceae: cladistics
and classification. Timber Press,
Portland, 752 pp.
Cates,
R.G. 1980. Feeding patterns of
monophagous, oligophagous, and
polyphagous insect herbivores:
the effect of resource abundance
and plant chemistry. Oecologia,
46: 22-31.
Coddington,
J.A., Griswold, C.E., Dávila,
D.S., Peñaranda, E. and Larcher,
S.F. 1991. Designing and testing
sampling protocols to estimate
biodiversity in tropical
ecosystems. In: E.C. Dudley
(Ed.), The unity of evolutionary
biology; proceedings of the
fourth international congress of
systematic and evolutionary
biology. Dioscorides Press,
Portland, Oregon, pp. 10-48.
Colwell,
R.K. and J.A. Coddington. 1994.
Estimating terrestrial
biodiversity through
extrapolation. Phil. Trans. R.
Soc. Lond. B, 345: 101-118.
Crutwell,
R.E. 1974. Insects and mites
attacking Eupatorium odoratum in
the Neotropics 4. An annotated
list of the insects and mites
recorded from Eupatorium odoratum
L., with a key to the types of
damage found in Trinidad. Tech.
Bull. Commmonw. Inst. Biol.
Contr., 17: 87-125.
Farrell,
B.D., Mitter, C. and Futuyma,
D.J. 1992. Diversification at the
insect-plant interface.
Bioscience, 42: 34-49.
Gagné,
R.J. 1994. The gall midges of the
Neotropical region. Cornell
University Press, Ithaca.
Kremen,
C. 1994. Biological inventory
using target taxa: a case study
of the butterflies of Madagascar.
Ecol. Applic., 4: 407-422.
Kremen,
C., Colwell, R.K., Erwin, T.L.,
Murphy, D.D., Noss, R. and
Sanjayan, M.A. 1993. Terrestrial
arthropod assemblages: their use
in conservation planning.
Conserv. Biol., 7: 796-808.
Lawton,
J.H., Lewinsohn, T.M. and
Compton, S.G. 1993. Patterns of
diversity for the insect
herbivores on bracken. In: R.E.
Ricklefs and D. Schluter (Eds.),
Species Diversity in Ecological
Communities. University of
Chicago Press, Chicago, pp.
178-184.
Lewinsohn,
T.M. 1991. Insects in flower
heads of Asteraceae in Southeast
Brazil: a case study on tropical
species richness. In: P.W. Price,
T.M. Lewinsohn, G.W. Fernandes
and W.W. Benson (Eds.),
Plant-Animal Interactions. John
Wiley, New York, pp. 525-559.
Lewinsohn,
T.M., and Prado, P.I.K.L. in
press. Padrões de associação
de insetos fitófagos com
Compostas (Asteraceae) na Serra
do Cipó. In: G.W. Fernandes
(Ed.), Serra do Cipó: Ecologia e
Evolução.
Lewinsohn,
T.M., Prado, P.I.K.L. and
Almeida, A.M. (manuscript).
Resource-centered inventories of
phytophagous insects.
Louda,
S.M. 1982. Distribution ecology:
variation in plant recruitment
over a gradient in relation to
insect seed predation. Ecol.
Monogr., 52: 25-41.
Müller-Schärer,
H., Lewinsohn, T.M. and Lawton,
J.H. 1991. Searching for weed
biocontrol agents - when to move
on? Biocontrol Science and
Technology, 1: 271-280.
Pimm,
S.L. 1991. The balance of nature?
Ecological issues in the
conservation of species and
communities. University of
Chicago Press, Chicago, 434 pp.
Polis,
G.A. and Winemiller, K.O. (Eds.).
1996. Food webs: integration of
patterns & dynamics. Chapman
and Hall, New York, 472 pp.
Prado,
P.I.K.L. and Lewinsohn, T.M.
1994. Genus Tomoplagia (Diptera,
Tephritidae) in the Serra do
Cipó, MG, Brazil: host ranges
and notes of taxonomic interest.
Rev. Bras. Ent., 38: 669-680.
PROBIO-MMA.
1998. Ações prioritárias para
a conservação da biodiversidade
do Cerrado e Pantanal. Workshop
(FUNATURA, Conservation
International, Universidade de
Brasília, Biodiversitas).
Brasília, março 1998.
Documentos preparatórios:
www.bdt.org.br/bdt/workcerrado.
Ricklefs,
R.E., and Schluter, D. (Eds.).
1993. Species diversity in
ecological communities:
historical and geographical
perspectives. University of
Chicago Press, Chicago, pp.
Ricklefs,
R.E., Kalin-Arroyo, M.T., Latham,
R.E., Lewinsohn, T.M., Lodge,
J.S., Platnick, N.I. and Wright,
D.S. 1995. The distribution of
biodiversity. In: V.H. Heywood
(Ed.), Global Biodiversity
Assessment. Cambridge University
Press, Cambridge, pp. 139-173.
Root,
R.B. and Cappuccino, N. 1992.
Patterns in population change and
the organization of the insect
community associated with
goldenrod. Ecol. Monogr., 62:
393-420.
São
Paulo. Secretaria de Estado do
Meio Ambiente. 1997. Cerrado:
bases para a conservação e uso
sustentável do estado de São
Paulo. Secretaria do Meio
Ambiente, São Paulo, 113 pp.
Thompson,
J.N. 1997. Conserving interaction
biodiversity. In: S.T.A. Pickett,
R.S. Ostfeld, M. Shachak and G.E.
Likens (Eds.), The ecological
basis of conservation:
heterogeneity, ecosystems, and
biodiversity. Chapman and Hall,
New York, pp. 285-293.
Thompson,
J.N. 1994. The coevolutionary
process. University of Chicago
Press, Chicago, 376 pp.
Zwölfer,
H. 1988. Evolutionary and
ecological relationships of the
insect fauna of thistles. Ann.
Rev. Entomol., 33: 103-122.
|